DNA損傷和修復----阿拉丁試劑

DNA損傷和修復的機制
細胞DNA的損傷與突變和癌癥的發生有關。人類細胞中的DNA每天要經歷數千到數百萬次的破壞事件,這些事件是由外部(外源性)和內部代謝(內源性)過程產生的。細胞基因組的改變會在DNA轉錄中產生錯誤并隨后翻譯成信號傳遞和細胞功能所必需的蛋白質。如果突變在有絲分裂之前沒有修復,基因組突變也可以傳遞到子代細胞。一旦細胞失去有效修復受損DNA的能力,就有三種可能的反應(圖1)。
1. 細胞可能會衰老,即不可逆地休眠。2005年,多個實驗室報告說,在體內和體外,癌細胞都可能發生衰老,阻止有絲分裂,并阻止細胞進一步進化。
2. 細胞可能發生凋亡。足夠的DNA損傷可能觸發凋亡信號級聯,迫使細胞進入程序性細胞死亡。
3. 細胞可能變成惡性的,也就是說,發展出不朽的特征并開始不受控制的分裂。
圖1所示。導致衰老、細胞凋亡或癌癥的細胞DNA損傷和修復的途徑
為了補償DNA損傷的程度和類型,細胞已經發展出多種修復過程,包括錯配、堿基切除和核苷酸切除修復機制,幾乎沒有過程冗余。如果發生了壓倒性的損傷,而不是消耗能量來有效地修復損傷,細胞可能已經進化到凋亡或衰老。細胞修復的速度取決于細胞類型和細胞年齡等因素。
DNA損傷的來源
多年來,外源性損傷被認為是DNA突變導致癌癥的主要原因。然而,Jackson和Loeb提出,DNA損傷的內源性來源也對導致惡性腫瘤的突變起著重要作用環境和細胞來源都可能導致類似類型的DNA損傷。
DNA可以被物理和化學誘變劑攻擊。物理誘變劑主要是輻射源,包括來自太陽的紫外線(200-300納米波長)輻射。紫外線輻射產生共價鍵,交聯相鄰的嘧啶(胞嘧啶和胸腺嘧啶)堿基在DNA鏈。電離輻射(x射線)通過在細胞內產生自由基引發DNA突變,自由基產生活性氧(ROS),并導致雙螺旋中的單鏈和雙鏈斷裂。化學誘變劑可以將烷基共價連接到DNA堿基上;能使DNA堿基甲基化或乙基化的氮芥化合物是DNA烷基化劑的例子。致癌物原是化學惰性前體,經代謝轉化為高活性致癌物。這些致癌物可以與DNA反應形成DNA加合物,即附著在DNA上的化學實體。苯并[a]芘是一種多芳雜環,本身并不致癌。它在細胞色素P450酶的介導下經歷了兩次連續的氧化反應,產生苯并[a]芘二醇環氧化物(BPDE),這種致癌代謝物能夠形成共價DNA加合物(圖2)。
圖2。苯并[a]芘被P450酶氧化生成具有高度致癌性的苯并[a]芘二烯烴。
DNA損傷也可由內源性代謝和生化反應引起,其中一些尚不清楚水解反應可以部分或完全將核苷酸堿基從DNA鏈上裂解。連接嘌呤堿基(腺嘌呤或鳥嘌呤)和脫氧核糖磷酸鏈的化學鍵可以在被稱為脫嘌呤的過程中自發斷裂。據估計,哺乳動物細胞中每天發生10,000次脫嘌呤事件脫嘧啶(從胸腺嘧啶或胞嘧啶中失去嘧啶基)也會發生,但其速率比脫嘌呤低20至100倍。
脫氨發生在細胞內,腺嘌呤、鳥嘌呤和胞嘧啶環的胺基丟失,分別產生次黃嘌呤、黃嘌呤和尿嘧啶。DNA修復酶能夠識別和糾正這些不自然的堿基。然而,在隨后的DNA復制過程中,未經糾正的尿嘧啶堿基可能被誤讀為胸腺嘧啶,并產生C→T點突變。
DNA甲基化,一種特殊形式的烷基化,發生在細胞內,由于與s -腺苷蛋氨酸(SAM)的反應。SAM是一種含有高活性甲基的細胞內代謝中間體。在哺乳動物細胞中,甲基化發生在距離鳥苷基(G) 5′的胞嘧啶堿基(C)胞嘧啶環的5位,即序列CpG。突變誤差的一個重要來源是甲基化的5-甲基胞嘧啶產物的自發脫氨。胺基的損失導致胸腺嘧啶堿基,它不被DNA修復酶檢測到作為一個非自然的堿基。由此產生的取代保留在DNA復制中,產生C→T點突變(圖3)。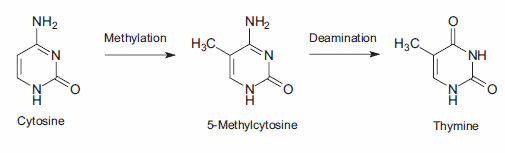
圖3。胞嘧啶的2期突變導致胸腺嘧啶,產生C→T點突變。
正常的代謝過程產生活性氧(ROS),通過氧化修飾堿基。嘌呤和嘧啶堿都容易氧化。最常見的突變是鳥嘌呤氧化為8-oxo-7,8-二氫鳥嘌呤,導致核苷酸8-oxo-脫氧鳥嘌呤sin (8-oxo-dG)。8-oxo-dG能夠與脫氧腺苷堿基配對,而不是像預期的那樣與脫氧胞苷堿基配對。如果這個錯誤沒有被錯配修復酶檢測到并糾正,隨后復制的DNA將包含C→a點突變。ROS還可能導致脫氧核糖核酸的脫嘌呤、脫嘧啶和單鏈或雙鏈斷裂。
在細胞周期S期的DNA復制過程中可能引入其他基因組突變。復制模板DNA的聚合酶有一個小但顯著的錯誤率,并且可能包含基于沃森-克里克配對的與模板DNA不正確的核苷酸。化學改變的核苷酸前體可以被聚合酶合并到生成的DNA中,而不是正常的堿基。此外,當復制含有大量重復核苷酸或重復序列的DNA片段(微衛星區)時,聚合酶容易“口吃”。這種酶的“口吃”是由于鏈滑移,當模板和復制的DNA鏈脫離正確的排列時。結果,聚合酶無法插入模板DNA所指示的正確數量的核苷酸,導致子鏈中的核苷酸過少或過多。
DNA可能發生單鏈和雙鏈裂解。單鏈斷裂可能是由于脫氧核糖基磷酸鏈的脫氧核糖部分受損造成的。斷裂也是ap -核酸內切酶去除磷酸脫氧核糖后堿基切除修復途徑的中間步驟。當單鏈斷裂發生時,核苷酸堿基和脫氧核糖主干都從DNA結構中丟失。雙鏈裂解最常發生在細胞經過s期時,因為DNA在展開作為復制模板時更容易斷裂。
DNA修復機制
雖然細胞能夠進化到凋亡或衰老狀態,但這些行動是最 后的手段。對于每一種類型的DNA損傷,細胞已經進化出一種修復損傷或消除破壞性化合物的特定方法。
o6 -甲基鳥嘌呤DNA甲基轉移酶(MGMT;DNA烷基轉移酶)根據DNA結構從鳥嘌呤中分離甲基和乙基加合物。該反應不是催化(酶促)反應,而是化學計量(化學)反應,每去除一個加合物就消耗一個MGMT分子。經過過表達MGMT的細胞對癌癥的抵抗力更強,可能是因為它們能夠抵消大量的烷基化損傷。Niture等人最近的一項研究報告稱,使用半胱氨酸/谷胱甘肽增強藥物和天然抗氧化劑可增加MGMT的表達。
DNA聚合酶,如聚合酶-δ,含有校對活性,主要參與復制錯誤修復。當檢測到錯誤時,這些聚合酶就會停止DNA復制的過程,然后反向從子DNA鏈中移除核苷酸,直到不正確的核苷酸明顯消失,然后重新啟動正向復制過程。在Pold1基因的兩個拷貝中都有點突變的小鼠表現出DNA聚合酶-δ的校對活性的喪失,并且與具有野生型基因或單拷貝突變的小鼠相比,發生上皮性癌癥的幾率顯著更高。
一組被稱為錯配切除修復(MMR)酶的蛋白質能夠糾正DNA聚合酶校對活動未檢測到的復制錯誤。MMR酶從子DNA中去除一個不正確的核苷酸,并使用W-C配對和父DNA鏈作為正確的模板來修復鏈這對于微衛星區域復制過程中產生的錯誤尤其重要,因為DNA聚合酶的校對活動不會檢測到這些錯誤。在較小程度上,MMR酶還糾正了由DNA氧化或烷基化引起的各種堿基對異常。這些突變包括含有o6 -甲基鳥嘌呤和8-氧鳥嘌呤的修飾堿基對,以及致癌物和順鉑加合物。12,13人錯配切除修復基因MSH2和MLH1突變與遺傳性非息肉病性結直腸癌(HNPCC)綜合征相關。
堿基切除修復和核苷酸切除修復
堿基切除修復(BER)涉及多個酶切除和替換單個受損的核苷酸堿基。主要由BER酶修復的堿基修飾是內源性氧化和水解破壞的堿基修飾。DNA糖基化酶切斷核苷酸堿基和核糖之間的鍵,使DNA的核糖磷酸鏈完整,但產生無嘌呤或無嘧啶(AP)位點。8-Oxoguanine DNA糖基化酶I (Ogg1)去除7,8-二氫- 8-Oxoguanine (8-oxoG),這是活性氧產生的堿基突變之一。人類OGG1基因的多態性與肺癌和前列腺癌等各種癌癥的風險相關。尿嘧啶DNA糖基化酶,另一種BER酶,切除胞嘧啶脫氨的產物尿嘧啶,從而防止隨后的C→T點突變n -甲基嘌呤DNA糖基化酶(MPG)能夠去除多種修飾的嘌呤堿基。
DNA中由BER酶作用產生的AP位點,以及由脫嘧啶和脫嘌呤作用產生的AP位點,可通過AP-核酸內切酶1 (APE1)的作用進行修復。APE1將磷酸二酯鏈5′裂解到AP位點。DNA鏈含有一個3 ' -羥基和一個5 ' -堿性脫氧核糖磷酸。DNA聚合酶β (Polβ)根據相應的W-C配對插入正確的核苷酸,并通過其相關的ap裂解酶活性去除脫氧核糖磷酸。x射線修復交叉互補基團1 (XRCC1)的存在是與DNA連接酶III (LIG3)形成異二聚體所必需的。XRCC1作為一個支架蛋白,為Polβ提供一個非活性的結合位點,并在修復位點將Polβ和LIG3酶結合在一起Poly(adp -核糖)聚合酶(PARP-1)與XRCC1和Polβ相互作用,是BER途徑的必要組成部分。18,19修復的最 后一步由LIG3完成,它將替換核苷酸的脫氧核糖連接到脫氧核糖基磷酸主干。這一途徑被命名為“短補丁誤碼率”。
另一種被稱為“長補丁誤碼率”的途徑用最少2個核苷酸的長度替換一條核苷酸鏈。據報道,修復長度為10到12個核苷酸。21,22 Longpatch BER需要增殖細胞核抗原(PCNA)的存在,它作為重組酶的支架蛋白其他DNA聚合酶,可能是Polδ和Polε, 24用于產生寡核苷酸瓣。現有的核苷酸序列被皮瓣內切酶-1 (FEN1)去除。然后寡核苷酸被DNA連接酶I (LIG1)連接到DNA上,封閉斷裂并完成修復用于確定短補丁與長補丁誤碼率路徑選擇的過程仍在研究中(圖4)。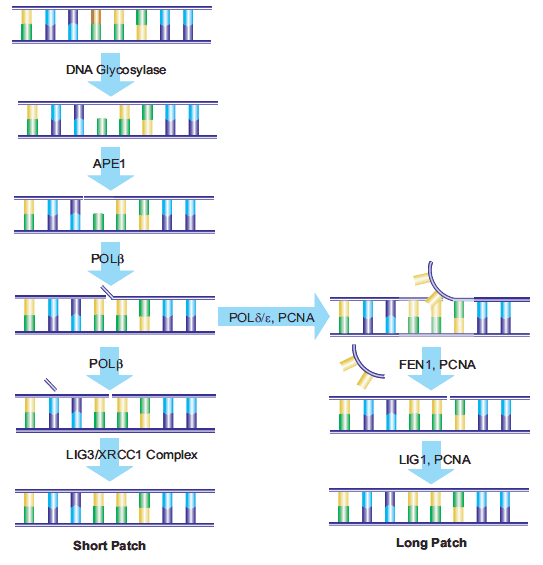
圖4。短補丁和長補丁誤碼率路徑示意圖。
雖然誤碼率可能通過長補丁途徑取代多個核苷酸,但短補丁和長補丁誤碼率的起始事件都是對單個核苷酸的損傷,從而對DNA雙螺旋結構的影響最小。核苷酸切除修復(NER)修復至少含有2個堿基的核苷酸鏈的損傷,并造成DNA的結構扭曲。NER的作用是修復單鏈斷裂以及外源的一系列損傷,如笨重的DNA加合物和紫外線輻射。同樣的途徑也可以用來修復氧化應激造成的損傷在哺乳動物細胞中,有超過20種蛋白質參與NER通路,包括XPA、XPC-hHR23B、復制蛋白A (RPA)、轉錄因子TFIIH、XPB和XPD DNA解旋酶、ERCC1- xpf和XPG、Polδ、Polε、PCNA和復制因子c。切除修復交叉互補(ERCC1)基因的過表達與非小細胞肺癌細胞對順鉑的耐藥有關29,并對應于增強的DNA修復能力30全 球基因組NER (GGR)修復整個基因組的損傷,而一種稱為轉錄偶聯修復(TCR)的特定NER通路在活性RNA聚合酶轉錄期間修復基因。
雙股斷裂的修復
DNA中的雙鏈斷裂會導致基因組序列的丟失和重排。這些斷裂可以通過非同源端連接(NHEJ)或同源重組(HR)進行修復,也稱為重組修復或模板輔助修復。
當細胞處于S/G2晚期且模板最近被復制時,HR通路被激活。這種機制需要存在一個相同或幾乎相同的序列,通過著絲粒連接到受損的DNA區域,作為修復模板。由該機制修復的雙鏈斷裂通常是由復制機制試圖通過單鏈斷裂或未修復的病變進行合成引起的,導致復制叉的崩潰。
當姐妹染色單體不能用作HR模板時,非同源末端連接(NHEJ)用于細胞周期的其他點。當這些斷裂發生時,細胞還沒有復制包含斷裂的DNA區域,因此與HR途徑不同,沒有相應的模板鏈可用。在NHEJ中,Ku異二聚體蛋白在沒有可用模板的情況下定位斷裂DNA鏈的兩端進行修復,在此過程中丟失序列信息。多個酶參與了再連接過程,包括DNA連接酶IV, XRCC4和DNA依賴蛋白激酶(DNA- pk)。32,33 NHEJ具有固有的誘變性,因為它依賴于兩個待連接DNA片段的單鏈尾部之間的偶然配對,稱為微同源性(圖5)。在高等真核生物中,NHEJ修復需要DNA- pk,無論是通過主要機制還是通過替代備份機制(D-NHEJ)。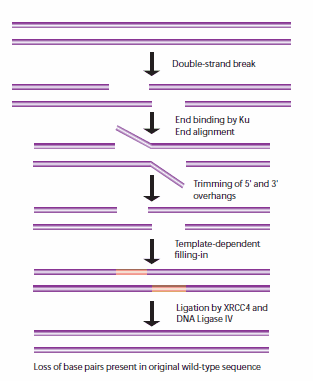
圖5。NHEJ修復DNA雙鏈斷裂的一般機制。
未來的應用
雖然DNA損傷是癌細胞發育和進化的關鍵因素,但持續的損傷被用作癌癥臨床治 療的一部分,迫使惡性細胞凋亡或衰老。許多化療藥物,如博萊霉素、絲裂霉素和順鉑,都是有效的,因為它們會進一步破壞癌細胞的DNA,而癌細胞的復制速度比周圍組織更快。細胞DNA修復機制是一把雙刃劍;通過減少可能導致癌癥的突變,這些過程努力保持基因組的完整性,但在惡性細胞中,同樣的機制允許這些細胞在額外的DNA損傷中幸存下來,并繼續不受控制地生長。為了阻斷癌細胞內的這種生存機制,目前正在進行臨床試驗,使用抑 制劑抑 制特定的DNA修復酶,包括MGMT、PARP和DNA- pk。35-38

標簽:DNA損傷和修復的機制
全部評論(0條)
推薦閱讀
-

- DNA損傷和修復----阿拉丁試劑
- 細胞DNA的損傷與突變和癌癥的發生有關。人類細胞中的DNA每天要經歷數千到數百萬次的破壞事件,這些事件是由外部(外源性)和內部代謝(內源性)過程產生的。
-

- 磁性納米顆粒在RNA和DNA分離中的應用----阿拉丁試劑
- 在過去的幾十年里,磁性納米顆粒(MNPs)越來越多地用于分離和區分生物分子,這也是目前大多數分子診斷程序的基礎。MNPs的大小、形態和分散性賦予了它們對生物分子的特異性、親和力和結合能力。
-

- 上新 | T4核酸內切酶V:引領高效DNA損傷修復!
- 在嘧啶二聚體的5’端切割糖苷鍵,從而釋放出二聚體并留下一個AP位點,隨后AP裂解酶活性通過 β 消除切割 AP 位點,與 3'-α、β-不飽和醛和5'-磷酸末端形成1個核苷酸DNA間隙
-
- 新品│Endonuclease VIII:精準切割DNA受損堿基,助力基因損傷修復研究
- 環介導等溫擴增(LAMP)高效快速,應用場景更廣泛
-
- 上新 | 核酸內切酶IV:翌圣DNA損傷修復酶家族又添一員,開啟高效修復新篇章!
- 識別DNA鏈中的脫嘌呤/脫嘧啶位點(AP位點),通過切割AP位點5'端的磷酸二酯鍵,產生3'-羥基和5'-脫氧核糖磷酸末端(dRP)
-

- 貼壁細胞培養指南:接種、增殖和收獲----阿拉丁試劑
- 細胞培養是一項艱難的工作。因為這是一個高度技術性的過程,當你從一個來源提取細胞并在另一個條件下去對他們進行操作時,可能會出現很多問題,比如受到外界的污染,或者生長速度非常緩慢。
-

- Fries重排----阿拉丁試劑
- Fries重排反應是指在路易斯酸或布朗斯特酸( 如HF、AlCl3, BF3, TiCl4, 或 SnCl4)催化下,以酚酯作為原料重排合成鄰位或對位酰基酚的反應[1]。
-

- 免疫沉淀技術----阿拉丁試劑
- 免疫沉淀(IP)是應用最廣泛的免疫化學技術之一。
-

- Heck反應----阿拉丁試劑
- Heck反應是有機化學中最為廣泛使用的碳-碳偶聯反應之一。Heck反應是烯烴與芳基或乙烯基鹵化物(或三氟甲磺酸酯)用于合成取代烯烴的鈀催化交叉偶聯反應。
-

- Diels-Alder反應----阿拉丁試劑
- Diels-Alder反應----阿拉丁試劑
-

- 卡爾費休庫侖法和容量法之間的區別----阿拉丁試劑
- 卡爾費休(KF)滴定法是一種具體的水分測定技術,被工業和科學家廣泛使用。根據其產生碘的方式不同,可以分為庫侖法和容量法兩種。
-

- 蛋白樣品制備流程----阿拉丁試劑
- 蛋白分析的第一步是蛋白提取,蛋白提取方法包括化學抽提法、物理抽提法和生物學抽提法。蛋白提取無論通過機械處理還是基于去垢劑的提取方法,都會不可避免地破壞細胞穩態,導致蛋白降解或不穩定。
-

- 代謝信號通路----阿拉丁試劑
- 細胞內穩態是由幾個關鍵代謝途徑的協調活動調節。這些過程包括碳水化合物代謝、脂質代謝、谷氨酰胺代謝和核苷酸代謝,它們維持細胞的能量狀態,并提供必要的成分以確保細胞正常功能。
-
- 白蛋白的應用----阿拉丁試劑
- 白蛋白是人體血漿中最主要的蛋白質,對維持血容量和體液平衡有重要作用。它是一種溶于水且遇熱凝固的球形單純蛋白,分子結構于1975年闡明,其為含585個氨基酸殘基的單鏈多肽,分子形狀呈橢圓形。
-

- 羥醛縮合反應----阿拉丁試劑
- 什么是羥醛縮合反應? 醛醇縮合反應是由Charles Wurtz提出的一種有機反應,他于1872年首次從乙醛中制備出β-羥基醛[1]。
-

- 生物緩沖液選擇指南----阿拉丁試劑
- 生物緩沖液選擇指南----阿拉丁試劑


①本文由儀器網入駐的作者或注冊的會員撰寫并發布,觀點僅代表作者本人,不代表儀器網立場。若內容侵犯到您的合法權益,請及時告訴,我們立即通知作者,并馬上刪除。
②凡本網注明"來源:儀器網"的所有作品,版權均屬于儀器網,轉載時須經本網同意,并請注明儀器網(www.ghhbs.com.cn)。
③本網轉載并注明來源的作品,目的在于傳遞更多信息,并不代表本網贊同其觀點或證實其內容的真實性,不承擔此類作品侵權行為的直接責任及連帶責任。其他媒體、網站或個人從本網轉載時,必須保留本網注明的作品來源,并自負版權等法律責任。
④若本站內容侵犯到您的合法權益,請及時告訴,我們馬上修改或刪除。郵箱:hezou_yiqi











參與評論
登錄后參與評論